
洋葱细胞质雄性不育系小孢子败育的细胞形态学结构
2017-05-23 23:22:25程雨宋策陈典王勇
江苏农业科学 2017年7期
程雨+宋策+陈典+王勇


摘要:为了探索洋葱细胞质雄性不育系小孢子败育的时间和形式,通过石蜡切片和电镜技术对小孢子的发育进行观察。结果表明,在花粉母细胞时期,不育系JA花药室壁较保持系厚且有凹凸现象;在四分体时期,小孢子不饱满呈月牙状,绒毡层与药室壁完全脱离,细胞质浓缩、空泡化;在小孢子发育时期,小孢子细胞质发生浓缩、降解、严重空泡化,绒毡层完全解体;在成熟花粉粒时期,小孢子完全干瘪聚集在一起,花药败育。由此推测,洋葱不育系JA小孢子发育异常始于花粉母细胞时期,在四分体时期呈现出败育,败育的原因在于绒毡层的提早解离。
关键词:洋葱;雄性不育;性状转育;四分体;小孢子;败育;细胞形态学结构;绒毡层;解离
中图分类号: S633.201文献标志码: A文章编号:1002-1302(2017)07-0107-04
洋葱(Allium cepa L.)为百合科葱属植物,别称圆葱、葱头,因其具有很好的食疗保健作用和极高的药用价值被誉为“菜中皇后”,在世界蔬菜生产面积上,排在马铃薯和番茄之后,位居第三[1]。我国是世界上洋葱的第一大生产国和出口国,产地主要分布在内蒙古、甘肃、云南、山东、黑龙江等地[2]。洋葱是最早育成并在生产上应用1代杂种的蔬菜作物之一[3]。洋葱杂种优势明显,自交衰退严重,其1代杂种一般能增产20%~50%,其应用价值已被生产实践所证实[4-5]。但洋葱为2年生异花授粉作物,其花器小,花期不集中,单花结籽率低,人工去雄成本高、难度大,因此利用雄性不育系是配制杂交种子的有效途径[6] 。研究洋葱细胞质雄性不育的发生机理对于 F1杂种的生产具有重要的作用。目前,洋葱细胞质雄性不育系主要分3类,即S型、T型和近年发现的瓣化型,其中S型在生产上应用广泛[7-9]。前人对洋葱CMS细胞学研究表明,洋葱花药败育大都与绒毡层发育有关,花粉母细胞没有异常表现。绒毡层的发育异常大致可归纳为3种形式:一是在四分体时期,绒毡层细胞提前解体,不能供给小孢子生长发育所需要的营养;二是在二分体时期,绒毡层过度肥大,而且会延迟解体,这2种类型中,绒毡层的解体总是发生在小孢子产生异常现象之前;三是绒毡层形态完全正常,只是存在时间过长[10-12]。一般研究认为,绒毡层细胞的异常死亡是造成小孢子败育的主要原因;但也有研究表明,不育系与保持系绒毡层细胞变化不大。连高山等对某一洋葱CMS研究发现,与可育系花粉母细胞相比,绒毡层细胞变化不大,但花粉母细胞次生壁和胼胝质合成受阻,细胞壁变薄,细胞内出现泡状结构,有些泡状体可相互融合,这种自噬现象的确与植物花药败育过程的细胞的败育或者说程序化死亡有关[13]。不同的雄性不育材料间小孢子败育发生的时期和原因并不完全一致,掌握败育发生的时期和成因对于雄性不育的转育和在生产上的应用意义重大。本试验对洋葱不育系JA及其保持系JB花粉发育过程进行观察分析,从形态学和细胞学上对洋葱细胞质雄性不育发生进行系统的跟踪和比较,确定其不育发生的时期,推测不育发生的原因,从而为该不育系发生的生理学和分子生物学研究提供参考,进而为该雄性不育性状的转育和利用提供理论指导。
1材料与方法
1.1试验材料
洋葱雄性不育材料JA和保持系JB,由东北农业大学园艺学院葱蒜课题组发现并选育。2013、2014年秋分别将JA和JB的种球种植于东北农业大学园艺站加温温室中,次年4月抽薹,开花后分别采集0.8~1.0、1.8~2.0、2.7~3.0、38~4.0 mm的花蕾,冰上剥离花药备用(经预试验确定其对应典型发育时期为花粉母细胞时期、二分体和四分体时期、单胞花粉粒时期、成熟花粉粒时期)。
1.2试验方法
1.2.1不育株与可育株的形态特征在播种期到开花期分别标记植株编号,定期到温室中观察不育系与保持系的形态特征并拍照记录。
1.2.2花药显微结构在开花盛期将采集的不同时期花药样本,放入FAA固定液中固定,石蜡切片法制片,切成厚 6 μm 的蜡带,用梅氏黏贴剂粘在载玻片上,1%番红染色,05%固绿复染,中性树胶封片,在显微镜下观察并照相。
1.2.3花药超微结构参考显微结构观察结果,将不同发育时期的花药放入2.5%戊二醛进行前固定,用0.1 mol/L磷酸缓冲液进行漂洗,在1%四氧锇酸固定液中固定2.5 h,再用0.1 mol/L磷酸缓冲液进行漂洗,经乙醇脱水后用1 ∶1比例的100%乙醇+100%丙酮脱水10 min,再用100%丙酮脱水10 min,固定及漂洗程序温度均保持在4 ℃,浸透包埋后在超薄切片机上将其修成50~60 nm的小块,用醋酸铀-枸橼酸铅双染色,最后在H-7650透射电镜下观察并照相。
2结果与分析
2.1不育株与可育株的形态特征
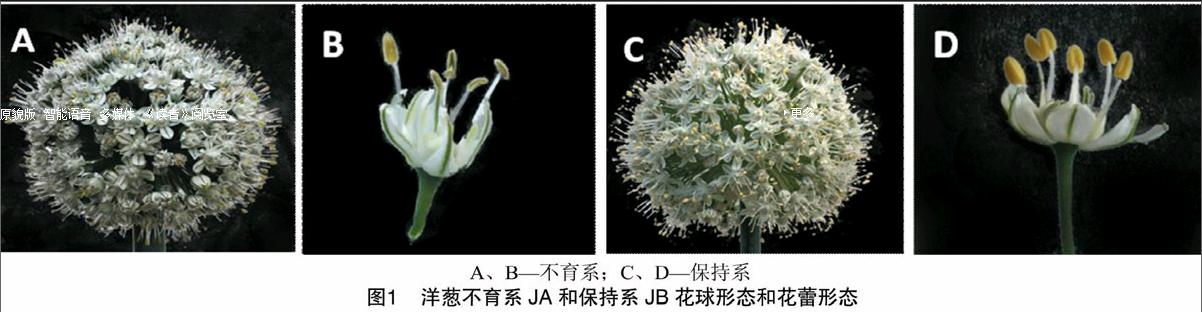
从幼苗期到开花期,洋葱不育株与可育株外形比较整齐一致,生长势无规律性差异,花球大小也无明显差异,但花蕾从表型上容易区分,即不育系JA花蕾虽然开放,但花丝逐渐缩小干枯,花药干瘪无花粉,多呈墨绿色后变褐色;保持系JB花蕾外形饱满,花丝发育正常,花药饱满,呈黄色(图1)。
2.2花药的显微结构
2.2.1花粉母细胞时期洋葱雄蕾原基分化发育成4个药室(图2-A、图2-B),每个药室的造孢细胞分化成若干个花粉母细胞。保持系JB花粉壁发育形状规则,由图2-C可以看出,花药壁由表皮、药室内壁、中层、绒毡层组成,中层细胞核清晰可见,绒毡层细胞核染色深,核较大排列在花粉母细胞周围。不育系JA花粉母细胞发育正常,但花粉囊细胞形状不规则,药室壁較保持系厚有凹凸现象,中层发育迟缓,花粉母细胞周围被绒毡层细胞紧紧包围(图2-D)。
2.2.2四分体时期保持系JB花粉母细胞经过2次减数分裂形成四分体,也有未完成减数分裂的二分体存在,胞壁染色较浅,药室表皮、内壁发育成熟,中层细胞排列整齐,细胞核清晰可见,绒毡层发育达到顶点,绒毡层细胞质浓郁发达,细胞核大、染色深(图2-E)。不育系JA花粉母细胞正常发育成四分体,但幼嫩小孢子没有保持系饱满,似月牙状(图2-F),药室表皮、内壁细胞膨大,比可育系厚,而中层细胞无发育迹象,细胞核染色较浅,绒毡层完全脱离药室壁,紧紧围绕四分体和二分体,胞质有明显的浓缩现象并空泡化。
2.2.3游离小孢子时期保持系JB花药中包围四分体的胼胝质解体,释放出小孢子,小孢子细胞核染色深清晰可见,绒毡层胞质降解,但其核染色仍较深,中层细胞没有明显变化,表皮和药室内壁高度发达,药室内壁出现纤维组织,细胞核染色较深(图2-G)。不育系JA小孢子从表面看染色透明類似泡状,中层细胞退化,绒毡层完全退化,看不到细胞核等结构,药室内壁形成粗大的纤维组织(图2-H)。
2.2.4成熟花粉粒时期保持系JB绒毡层在供应花粉发育所需要的营养后完全解离,整个花粉粒染色较深,形状饱满,花粉粒均匀地分散在药室内,相邻花粉囊相通,药室内壁纤维化细胞出现双核(图2-I)。不育系JA空瘪的花粉粒收缩挤在一起,粗大的纤维组织加厚,不具有开裂腔结构,药室严重收缩变形,内壁几乎贴在一起,纤维层不能形成裂口,花粉囊不相通(图2-J)。
2.3花药的超微结构
2.3.1四分体时期不育系JA绒毡层出现明显胞质收缩、染色质浓缩、泡状结构(图3-A )、内质网溢满、线粒体膨胀(图3-B),小孢子形状不均匀,细胞质出现浓缩并有少量泡状结构(图3-C),所以从形态上推测不育系JA绒毡层细胞在四分体时期之前就开始了细胞编程性死亡(PCD)。保持系JB小孢子正常发育,核质浓郁,没有浓缩现象(图3-D)。
2.3.2游离小孢子时期洋葱游离小孢子时期,保持系JB绒毡层释放胞质内容物帮助形成小孢子壁,出现降解现象(图3-E、图3-F),不育系JA绒毡层退化严重,虽然有类似小孢子壁结构但小孢子紧密聚集在一起,细胞器无法辨别,出现大量泡状结构,不能正常发育(图3-G)。
2.3.3成熟花粉粒时期保持系JB随着绒毡层细胞内容物的释放,绒毡层形成线状,但仍有残留(图3-H),花粉粒逐渐成熟,电子密度浓厚,染色很深(图3-I)。不育系JA绒毡层无法辨别,花粉壁形成,但小孢子干瘪、变形,内容物只剩残留,完全败育 (图3-J)。
3讨论与结论
前人研究表明,植物雄性不育小孢子败育在不同植物材料中发生时期有所差异,相同材料不同品种间也存在差异[14-17]。本研究中不育系JA在花粉母细胞时期,小孢子花粉母细胞正常发育,药室表皮和内壁细胞较保持系有加厚凹凸现象;在四分体时期出现败育迹象,小孢子不饱满呈月牙状,超微观察显示小孢子出现细胞质浓缩现象,绒毡层过度肥大,出现液泡,药室表皮和内壁细胞膨大;游离小孢子时期,小孢子细胞质浓缩加重,堆积在一起无法正常发育;成熟花粉粒时期,小孢子空瘪,孢子囊皱在一起,中层和绒毡层完全退化,药室变形不开裂。笔者分析比较发现,小孢子在四分体时期出现败育迹象;但是李园园等研究发现,洋葱细胞质雄性不育系63A四分体时期,四分体形成正常,而小孢子发育时期,小孢子细胞质发生浓缩、降解,绒毡层完全解体[12],这与杜敏霞等的研究结果[18]相似。而王瑞丽等研究发现,洋葱细胞质雄性不育系8A在花粉母细胞时期就发生了败育[19]。小孢子败育时期的差异可能是由材料不同造成的。
从形态学角度来看,大多数植物的小孢子败育都涉及到了绒毡层的发育异常。绒毡层作为花药壁最内层的特殊体细胞,与小孢子母细胞以及花药发育后期的小孢子直接相互作用,在花粉发育过程中起着至关重要的作用,绒毡层以程序性死亡(PCD)的形式适时为小孢子发育提供营养和结构物质,分泌胼胝质酶,参与花粉粒外壁的形成,同时也为花粉的成熟提供空间。当环境条件改变或调控绒毡层细胞 PCD 的基因发生突变时,都会导致绒毡层细胞降解的提前或延迟,致使花粉发育受阻,并最终形成雄性不育[20-22]。王婵等研究发现,大葱不育系58A花药绒毡层提前解体,影响了正常小孢子的形成,导致花粉败育[23]。李园园等也认为,洋葱不育系小孢子败育与绒毡层提前衰退有关[12],类似结论在辣椒[24]、油菜[25]、水稻[26]等作物上均有报道。洋葱雄性不育系JA在四分体时期,绒毡层细胞出现细胞质、染色质浓缩,内质网溢满,线粒体膨胀等程序性死亡现象,而其保持系四分体时期绒毡层发育到顶点,但未出现退化现象,直到游离小孢子时期绒毡层才退化分解,由此推断不育系绒毡层的提前解离是造成小孢子营养不足、逐渐凋亡,最终导致败育的原因之一。
形态学上的表现是基因表达的结果,本研究确定了洋葱不育系JA小孢子败育的发生时期和直接原因;但有关分子信息的研究还不明确,如哪些基因引起绒毡层提早解离,小孢子败育除了线粒体基因的表达,与核基因是否还存在着某种关联,洋葱的雄性不育发生机理的研究无论是对雄性不育的利用,还是对洋葱的杂交制种都具有重要意义,这也是下一步研究的方向。
参考文献:
[1]梁毅,王永勤,于春霞,等. 中国洋葱产业的回顾与展望[J]. 中国农学通报,2009,25(24):308-312.
[2]高金龙,张衍荣,郑锦荣,等. 我国洋葱产业发展的思考[J]. 广东农业科学,2009(6):282-284.
[3]王建军,侯喜林,宋慧,等. 洋葱育种研究进展[J]. 中国蔬菜,2003(4):57-59.
[4]谭其猛. 蔬菜育种[M]. 北京:农业出版社,1980:244-307.
[5]吴海涛,马蓉丽,刘洪炯,等. 洋葱细胞质雄性不育系选育研究进展[J]. 园艺学报,2009,36(2):297-302.
[6]李园园. 洋葱细胞质雄性不育的机理研究[D]. 南京:南京农业大学,2006.
[7]Jones H,Emsweller S. A male sterile onion[J]. Journal of the American Society for Horticultural Science,1936,34:582-585.
[8]Berniger. Contribution a letude dela sterility emalede loignon (Allium cepa L.)[J]. Ann Amelior Plants,1965,15:183-199.
[9]马有会,王火旭,崔成日,等. 洋葱瓣化型细胞质雄性不育系 psf-A的遗传分析[J]. 北方园艺,2011(6):23-24.
[10]Holford P,Croft J,Newbury H J.Structural studies of microsporogenesisin fertile and male-sterilr onions(Allium care L.) containing the CMS-Scytoplasm[J]. Theoretical and Applied Genetics,1991,82:745-755.
[11]Patil J A,Jadhav A S,Rane M S. Male-sterility in mahrashtra onion (Allium cepa L .)[J]. Research Journal of the Mahatma Phule Agricultura University,1973,40:29-31.
[12]李園园,杨清,严继勇,等. 洋葱63A细胞质雄性不育与绒毡层的提早衰退有关[J]. 作物学报,2006,32(3):369-372,475-476.
[13]连高山,王新宇. 洋葱雄性不育系花药在花粉母细胞时期的败育与自噬现象紧密相关[C]//中国细胞生物学学会2013年全国学术大会论文集. 武汉,2013.
[14]王兰兰,王晓林,魏兵强,等. 辣椒雄性不育系及保持系小孢子发育的细胞学比较[J]. 西北农业学报,2015,24(1):115-118.
[15]王开芳,张咏梅,张金文,等. 甘蓝型油菜细胞质雄性不育系105A花药败育的细胞学观察[J]. 中国农学通报,2015,31(13):76-80.
[16]郑洁,王有国,崔光芬,等. 东方百合可育品种及其不育突变体花药形成的细胞学观察[J]. 西南农业学报,2015,28(3):1246-1250.
[17]董庆华,利容千,王建波. 菜薹细胞质不育系小抱子发生的细胞形态学研究[J]. 园艺学报,1997,24(2):150-154.
[18]杜敏霞,刘湘萍. 洋葱雄性不育材料小孢子发生的细胞形态学观察[J]. 华北农学报,2007,22(增刊3):48-51.
[19]王瑞丽,马蓉丽,成妍,等. 洋葱细胞质雄性不育系及保持系小孢子发生的细胞学观察[J]. 中国农学通报,2016,32(1):43-46.
[20]Varnier A L,Mazeyrat-Clourbeyre F,Sangwan R S,et al. Programmed cell death progressively models the development of anther sporophytic tissues from the tapetum and is triggered in pollen grains during maturation[J]. Journal of Structural Biology,2005,152(2):118-128.
[21]张虹,梁婉琪,张大兵.花药绒毡层细胞程序性死亡研究进展[J]. 上海交通大学学报(农业科学版),2008,26(1):86-90.
[22] Dawson J,Wilson Z A,Aarts M G M. Microspore and pollen development in six male-sterile mutants of Arabidopsis thalian[J]. Can J Bot,1993,71:629-638.
[23]王婵,赵泓,刘倩纯,等. 大葱CMS 58不育系及保持系花药和花粉的显微观察[J]. 中国蔬菜,2014(10):29-34.
[24]吕晓菡,方献平,柴伟国,等. 辣椒胞质不育系与保持系花药的细胞学和蛋白质组学差异分析[J]. 浙江大学学报(农业与生命科学版),2015,41(1):44-55.
[25]鲁美宏,孙万仓,孔德晶,等. 白菜型冬油菜不育系LRCMS花器生理生化特性及其雄蕊发育特征研究[J]. 西北植物学报,2014,34(3):509-515.
[26]魏磊. 紫稻型细胞质雄性不育水稻樱香不育系的细胞学研究[J]. 农业科学与技术(英文版),2015,3(3):459-461.
猜你喜欢
园艺与种苗(2021年10期)2021-11-28 23:31:51
湖北农业科学(2021年19期)2021-11-15 07:01:38
云南农业大学学报(自然科学)(2021年3期)2021-06-11 04:19:38
中国种业(2018年8期)2018-08-28 06:40:46
广西植物(2016年10期)2016-11-11 06:51:39
西南农业学报(2016年4期)2016-05-17 05:41:45
种子科技(2015年8期)2015-05-24 16:14:56
安徽农业科学(2015年13期)2015-01-13 06:20:26
中国瓜菜(2014年3期)2014-04-29 00:44:03
中国蔬菜(2013年8期)2013-01-28 04:52:48
